原链接:链接
我们现代人在分类学上被称为智人
(Homo sapiens)
,也有一种流行的说法称我们自己为恐怖直立猿。这是因为自我们的祖先走出非洲以来,一路伴随着的,是由我们祖先自身造成的无尽的杀戮与无数物种的灭绝。这些被灭绝的物种不仅包括本身是欧洲、亚洲大陆、美洲、东南亚群岛等地的土著物种,更包括了在我们恐怖直立猿十多万年前走出非洲之前便离开非洲大陆去闯荡的“表亲”——直立人、尼安德特人、丹尼索瓦人等等。所以,对这些被悲剧地“抄家灭族”的物种来说,“恐怖直立猿”的称谓我们当之无愧。
我们之所以能够在走出非洲之后,便犹如降维打击般地走到哪就屠杀到哪,遇不到哪怕丝毫的反抗,是因为我们所具有的独特的优势:直立行走、超大脑容量、社会属性。这一系列的生理改变一步一步地如连锁反应般,又如蛛网一般相互影响,最终使我们能够拥有文明与智慧。
直立行走
假如你是一只无忧无虑的森林古猿,本住在东非原始森林里,林中有果又有树,生活乐无边;谁知那地球盖亚祂蛮横不留情,板块位移撕开了大裂谷。沟壑现,高山起,挡住了西方带着暖湿气流的信风。
由于极短时间内发生的这一剧变,原始森林没有了暖湿气流的温养,在越发干燥的环境中,逐渐破败,迅速地从森林退化为了草原。
原本安逸和谐生活着的你,体会到了什么是家道中落。短短数代“猿”的时间,你亲眼看见一颗颗巨木枯死、折倒、腐败,一“猿”高的荒草在巨木腐朽的尸体上生根发芽,丰富的营养让荒草长得格外茂盛。
在越发稀疏的林间,食物越来越难寻觅。原来触手可得的浆果已经消失殆尽,费点精力能够抓到的树栖小动物也不见踪影。你不得不小心翼翼地从所剩无几的树上下来,小心翼翼地踏出迈向大地的第一步。
这是你的一小步,也是猿类的一大步。
习惯了弓着身子的你猛然发现,茂盛的荒草挡住了你的视线,你不得不直起身子来看得更远。不似树梢间,你能够辗转腾挪,把捕食者“秀翻”;在草丛中,你只有直起身子,才能够看得更远,才能够远远地避开捕食者。
你看着身边那些艰难直起身子,或者没有办法长时间直立的大多数同伴一个个地丧命于血盆大口,心惊胆战地与更加习惯于直立的一小部分同伴躲在远处瑟瑟发抖。
好在,你们的孩子继承了你们的特点。一代代下去,总是那些更加适应于直立的猿类得到了活着繁衍后代的机会。
没有了物产丰富的果树,草根、灌木远远不能让你填饱肚子,你的目光逐渐落在了在远方惬意咀嚼着草叶的大型食草动物身上。
刚习惯直立行走不久的你,远没有顶级猎食者那完美的硬件条件,没有搜寻猎物的强大嗅觉与听觉,没有充满爆发力的肌肉与流线型的身形,你只能笨拙地向猎物跑去。

图1 猩猩vs人
然而,就连普通食草动物的爆发力也不是你能比的,甚至不等你靠近,它便一蹬腿,溜了。
你没有任何捕猎技巧,带着对食物的渴望,你只能傻乎乎地远远吊在猎物屁股后面追击。但是,渐渐地,你发现你的目标已经疲惫不堪,而你却没有感到任何不适;直到猎物坚持不住休克倒下,你也仅需要大口喘气而已。
你不知道的是,后世有人对此进行了研究,将两足行走的志愿者和四足爬行的猩猩进行对比。发现相同情况下,两足行走所消耗的氧气和能量仅是四足爬行的四分之一[2]。
因环境剧变而被迫站起来的古猿却因此获得其他物种难以想象能力——极致的耐力,这使得并没有其他突出优势的古猿具有了独特的捕猎方式。
每一次细微的突变,微小的生存优势,在几十代乃至数百代的演化之后,更多有利于长途跋涉的表型被固定了下来:发达的足弓、体毛的消失、全身发达的汗腺、向两边导流汗液的眉毛。
草原上的食草动物与捕食者都是在几千万年间的相互影响中,形成以爆发力为主的选择压力。因此,耐力特长古猿的马拉松式的捕猎方式能够使其轻松地获得充足的能量。这不仅给沿袭于林间生活的群体内社交留出了充足的时间,影响其后相关基因频率的改变;更给高耗能的大脑的容量增加提供了能量基础。
脑容量增加
大脑作为最耗能的器官,仅占体重的2%,但却会消耗全身近25%的能量与氧气,需要无比丰富的血管网络进行供给。
这对于其他动物来说是难以承受的消耗。
对于捕食者来说,他们获得生存所需能量依靠的是隐匿身形的外表、极致爆发的肌肉以及撕开猎物的尖牙利齿。就算在无数的基因突变中,出现了那么一个稍微聪明的个体,等待它的也不会是明显的生存优势,而是在大脑剧增的耗能需求下,疲于奔命,最终饥饿而死。
对于食草动物来说同样如此,本就依靠不含多少能量的植物枝叶而生,身体一大半的部位都被包含多个胃的巨大的消化道所占据,获得能量的效率远远无法支撑一个高耗能的大脑。
然而,独特的捕猎方式给我们的祖先带来了充分的营养摄入,解开了对大脑能耗的限制,使得脑容量变大的突变不再有害。
充足的能量能够使其支撑一个聪明的大脑所需的消耗,而这个时候,智商的优势便能够显现出来,并最终在生存的压力下脱颖而出。
当然,大自然中的任何变化都不会是由单一因素造成的。
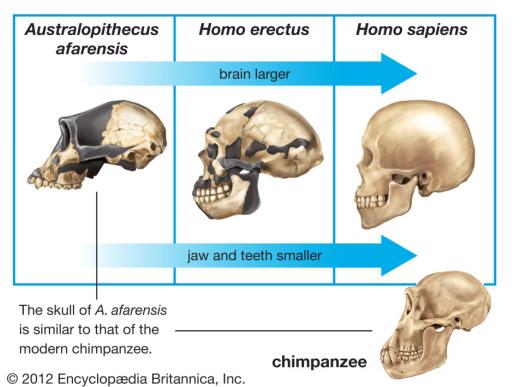
图2 脑容量的进化
短短数代,古猿从树冠来到平地。以往只需要处理眼前十来米范围内的信息,而在一望无际的草原,大脑需要处理的视觉信息瞬间增加好几个数量级,这对大脑处理信息的能力是一个不小的挑战。
同时,直立的行走方式也对大脑的平衡感控制产生了新的要求。
因直立行走而解放的双手带来了更多的感觉信息,物体的外形、大小、粗糙程度等等信息都需要一个精妙的大脑来处理。
这里并不是说有这个需求,大脑便会定向地向大脑容量的方向进化。而是在无数的没有方向的随机突变中,这些基因突变大多都是有害的,产生了那么少数几个能够使个体拥有更大脑容量的基因突变。
而这些具有些微优势的少数个体,在大脑能量需求能够得到满足的情况下,往往会具有更强的信息处理能力、更加协调的运动能力以及更加强大的学习能力。
这使得他们在同类中脱颖而出,在环境压力的选择下,能够留下更多的后代。当数百上千世代过后,整个族群的平均脑容量便会得到增加——变得更聪明。
脑容量的增加使得我们除了满足最基础的生存以外,还形成了更加丰富的社交方式。
但事物都有两面性,更大的脑容量意味着更大的大脑。婴儿的大脑需要在出生前就具有一定的功能以维持生存的基本需要,而直立行走的生活方式却限制了女性产道的大小。
在这样的生育困境下,我们依然保持着直立行走与硕大的高耗能大脑。这说明,在严酷的环境压力下,保持直立行走与聪明的大脑带来的适应性优势要超过生育困境所带来的危害。
不过,越发艰难的生育与哺育过程也促使着我们祖先的社会结构发生里程碑式的改变,这一改变,为我们的终极形态——文明的萌芽奠定了坚实的基础。
生育困境与社会属性
为了在保持充足脑容量和直立行走的情况下顺利产下后代,人类逐渐演化出三个策略:增强产道收缩能力以强行挤出胎儿、骨缝尚未连接且具有一定形变能力的胎儿头骨以及在大脑发育完全之前便将胎儿生出来。
然而,这也就造成出生的婴儿在初期十分脆弱,相对于其他动物来说,人类的婴儿其实都算是“早产儿”。
理查德·道金斯《自私的基因》一书中指出,个体只是基因的载体,是其延续自身的工具。也就是说,个体的一切行为(文明诞生以前),其最终目的都是基因的延续和繁衍。
对于女性来说,自己产下的婴儿可以肯定是自己的后代,其体内所含的基因也必定有一半来自自己。因此,消耗巨大精力,尽心尽力抚养其长大直至性成熟便有了充足的动力,
但由于这种远超其他动物的,难以想象的抚养难度,女性无力一人承担抚育后代的任务。
在蒙昧的原始时代,男性无法确定与自己交配的女性产下的后代一定属于自己。因此,这时候的男性与其他动物类似地选择了广撒网的“渣男”式繁衍方式。
这种情况下,女性便需要一些特别的方式来将男性留在身边,使其帮助自己抚养后代。
在漫长的两性博弈过程中,女性逐渐发展出隐蔽排卵与持续发情两大策略。
持续发情:一般其他动物为了能够留有充足的时间与精力去获取食物,往往会将繁衍后代的工作集中在一段时间内,这段时间也叫发情期,大多是在气候温暖、物资丰富的春天。人类高效的捕猎方式使得自身无需受到时节的限制;同时,发达的大脑能够从交配行为中得到更强烈的正反馈。为了应对生育困境,牢牢栓住男性,在长期的博弈过程中,女性的身体做出了巨大改变,延长发情期,使男性能够有更长时间留在女性身边;随着时间推移,逐渐将每一年的每一天都占满。可以说,人类并没有发情期的存在,或者说每时每刻都处于发情期中。
隐蔽排卵:然而,单单持续发情并不能使男性心甘情愿为女性照顾下一代。女性逐渐发展出第二个策略,隐蔽排卵。对于大多数动物来说,发情期便是排卵期,但人类女性的排卵期并不会在身体上表现出来。这样,“渣男”式的繁衍方式便不能保证男性自己的基因得到有效的传承。
在女性以上两大策略下,对于男性来说,为了保证自己基因的延续,最好的选择便是守株待兔,守住一个女性。同时,长期的共同生活也保证了没有外来男性的干扰,增加其对自己孩子的辨识度。在能够确认孩子是属于自己的后代后,父爱便有了萌芽的土壤,男性开始心甘情愿养育下一代。
母爱、爱情、父爱开始出现,家庭的纽带逐渐形成,而老人的存在也开始变得有意义。丰富的家族内情感交流与族群内的社交给人类带来了超乎想象的凝聚力,大规模的合作开始成为可能,文明萌芽的基础也开始形成。
我们之所以能够席卷全球,最重要的原因还是我们强大的社会属性,而这一社会属性又源自于家庭纽带催生出的共情心理。回过头看一看这一切,其根源竟然在于几百万年前的直立行走。直立行走使我们获得长途奔袭的能力,高效狩猎获得的充足营养又给我们脑容量变大奠定了基础,但更大的脑袋却使女性面临严峻的生育困境。在长期的性别博弈过程中,女性演化出隐蔽排卵和持续发情的生理特征,而这也促使了家庭的形成。这样的环境下,足够聪明的大脑逐渐在语言、沟通交流方面得到强化,具有同理心的利他行为开始出现。
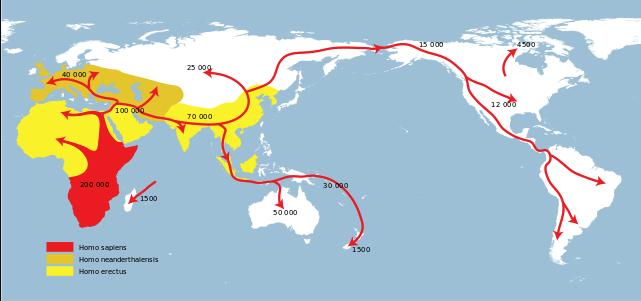
图 3.智人走出非洲[8](红色:智人;棕色:尼安德特人;黄色:直立人)
以上变化看起来轻描淡写,但其实是极其缓慢的演化过程,一些细微的新性状往往需要几万年乃至几十万年的时间来扩散至整个族群。
最初我们可能和非洲大草原上其他动物一样,随着季节追赶着雨水和食物,年复一年来回迁徙。但强大的长途跋涉能力使得人类能够走出舒适圈,前往更远的地方探索栖息地;聪明大脑的好奇心给了人类往外探索的动力。
从300万年前直立人第一次走出非洲开始,由最初共同祖先演化而来的,不同分支的古人类开始在其他大陆上蔓延,直到数万年前我们的祖先,智人席卷全球。
任何理论都需要有充分的证据来证实,但我们不可能乘坐时光机回到当初那个茹毛饮血的时代,跟着我们的祖先一步一步地观察他们的历史。
我们只能通过古代遗址的发现,并对其中遗留的信息进行深入地挖掘,才能够逐渐揭开人类发展的历史。
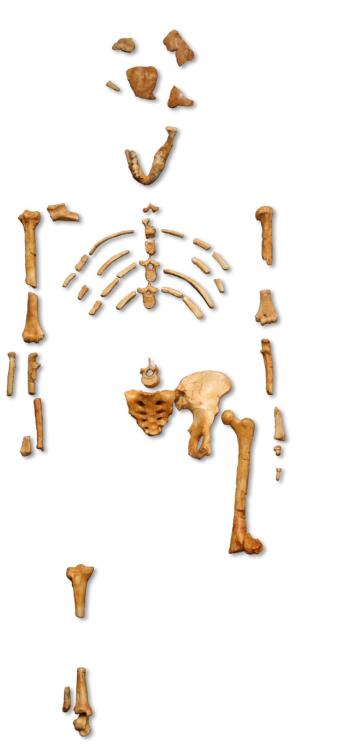
图 4. “露西”[8]
1974年,南方古猿“露西”较为完整的骨架在埃塞俄比亚被发现,并在之后被准确定位至318万-340万年前[3]。“露西”的发现是古人类学的一个里程碑,为古人类学研究提供大量线索与证据,自此古人类研究翻开了一个新的篇章。“露西”的骨骼特征表明她可直立行走,但脑容量不高,这说明脑容量的增加是在直立行走之后发生的。随着更多古遗迹的发现以及基因组学研究方法的进步,关于脑容量的证据与线索也越来越多地得以被发现[4]。
追溯人类的历史,进化树上的其他分支也是不可忽视的一部分。23万年到3万年前主要生活在欧洲北部寒冷地带的尼安德特人,30万年到3万年前生活在东亚北部的丹尼索瓦人,以及其他更多地区的土著人种的历史,都在越来越多遗址被发现的过程中被追根溯源[5]。
我们中国华夏民族自身的溯源研究,也在科研人员的不懈努力下,逐渐深入。现在,得益于古DNA提取技术的发展,古人类学工作者已经能够从数千乃至几万年前的骨骼样本中,在混杂的诸多微生物基因组里精确分辨出样本的数据。并运用生物信息、群体遗传学等知识,对其进行专业的统计学分析,追踪我们炎黄子孙的演化历史[6]。
技术的进步,使得我们从之前只能针对骨骼形态进行研究,到现在已经能够提取古DNA进行基因组层面的比较研究,人类历史的画卷正在不断清晰;各个不同地区人群的演化历史也渐渐深入,填补上这缺失的几块拼图。在将来,人类的源流图谱会更加清晰。
参考文献
[1] 史钧. 疯狂人类进化史[M]. 重庆出版社, 2016.
[2] Sockol M D, Raichlen D A, Pontzer H. Chimpanzee locomotor energetics and the origin of human bipedalism[J]. Proceedings of the National Academy of ences of the United States of America, 2007, 104(30):12265-12269.
[3] Kimbel W H, Delezene L K. "Lucy" Redux: A Review of Research on Australopithecus Afarensis[J]. American Journal of Physical Anthropology, 2009, 140 Suppl 49(S49):2-48.
[4] Leigh S R. Cranial capacity evolution in Homo erectus and early Homo sapiens.[J]. American Journal of Physical Anthropology, 2010, 87(1):1-13.
[5]Noonan J P, Coop G, Kudaravalli S, et al. Sequencing and analysis of Neanderthal genomic DNA[J]. science, 2006, 314(5802): 1113-1118.
[6] Yang M A, Fan X, Sun B, et al. Ancient DNA indicates human population shifts and admixture in northern and southern China[J]. Science, 2020.
[7] Human Evolution, 不列颠百科全书.
[8] 维基百科.
最后于 2023-12-15
被一只雄性灵长类动物编辑
,原因: 1145141919810